
Endozytosemechanismen die zentrale Rolle bei Antigenpräsentierung
Waffenwahl des Immunsystems
-
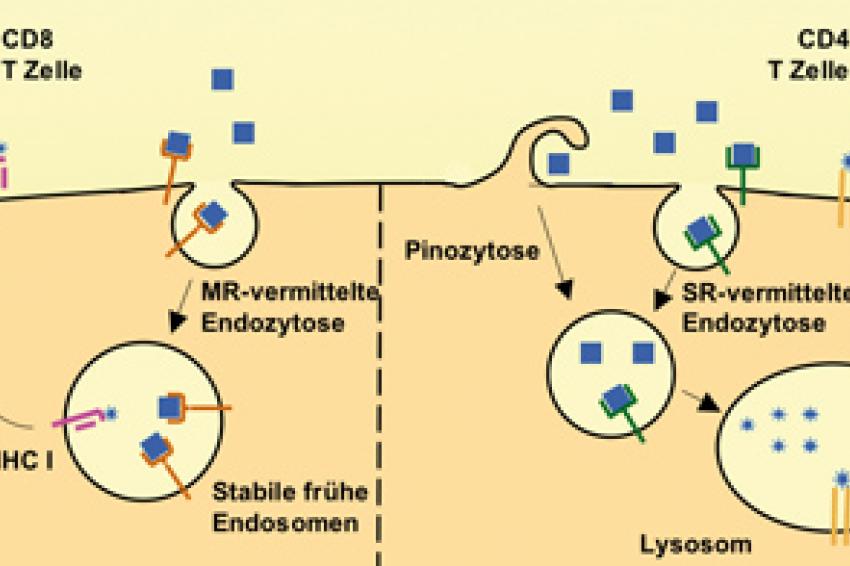 Abb. 1: Modell der Antigenpräsentation. Antigen wird simultan über mehrere Mechanismen in verschiedene Zellorganellen aufgenommen. Eines dient der MHC I Beladung zur Aktivierung von CD8+ T Zellen (links), das andere der MHC II Beladung zur Aktivierung von CD4+ T Zellen (rechts).
Abb. 1: Modell der Antigenpräsentation. Antigen wird simultan über mehrere Mechanismen in verschiedene Zellorganellen aufgenommen. Eines dient der MHC I Beladung zur Aktivierung von CD8+ T Zellen (links), das andere der MHC II Beladung zur Aktivierung von CD4+ T Zellen (rechts).
Zur Bekämpfung von Krankheitserregern oder Tumoren kann das Immunsystem gezielt CD4+ T Helfer oder CD8+ T Effektor Lymphozyten einsetzen. Wir konnten kürzlich zeigen, wie zwischen der Aktivierung dieser beiden Zelltypen gewählt wird. Hierzu setzen Antigenpräsentierende Zellen gezielt Endozytosemechanismen ein, um Pathogenkomponenten in separate Organellen aufzunehmen und so aufzubereiten, dass der korrekte T Zelltyp aktiviert wird. Dies eröffnet neue Möglichkeiten zur Impfstoff-Entwicklung.
Das Immunsystem setzt unterschiedliche Waffen gegen die verschiedenen Klassen von Krankheitserregern und Tumoren ein. T Lymphozyten („T Zellen") spielen hierbei eine wichtige Rolle als Regulatoren und Effektoren der Immunabwehr. Zytotoxische CD8+ T Zellen zerstören Virus-infizierte Zellen oder Tumorzellen, während die CD4+ T-Helfer Zellen andere Immuneffektoren steuern, wie zum Beispiel Makrophagen, die etwa in der Abwehr von Bakterien wichtig sind, oder B Zellen, die Antikörper produzieren oder die bereits erwähnten zytotoxischen CD8+ T Zellen gegen Viren. Grundsätzlich müssen T Zellen aktiviert werden, um ihre Funktion ausführen zu können. Dies ist die Hauptaufgabe der dendritischen Zellen, die deswegen auch als „professionelle" Antigen-Präsentierende-Zellen (APZ) bezeichnet werden. APZ nehmen Komponenten von Krankheitserregern (Antigene) auf, prozessieren diese und präsentieren sie schließlich den T-Zellen. Die für ein bestimmtes Antigen spezifischen T-Zellen werden dadurch aktiviert, teilen sich vielmals und können daraufhin ihre Effektor- oder Regulatorfunktionen verrichten. APZ haben verschiedene Möglichkeiten, Antigene aufzunehmen und den anderen Immunzellen zu präsentieren. Ein spezifischer Weg ist die rezeptorvermittelte Endozytose, in der Antigene über bestimmte Rezeptoren aufgenommen werden. Außerdem gibt es noch die Phagozytose, bei der große Partikel, wie etwa ganze Bakterien aufgenommen werden, und die Pinozytose, in der unspezifisch eine kleine Menge extrazelluläre Flüssigkeit mit allen darin enthaltenen Molekülen aufgenommen wird. Derartig aufgenommene Antigene werden von APZ degradiert und als Peptide an sog Major HistoCompatibility (MHC) Moleküle gebunden.
Diese werden an die Zelloberfläche befördert, um an T-Zellen präsentiert zu werden. CD8+ T Zellen nehmen Antigen auf MHC Klasse I Molekülen wahr, während CD4+ T Zellen Peptidbindung an MHC Klasse II Moleküle verlangen. Neben Antigenen, die extrazellulär aufgenommen werden, können auch endogene Antigene, also Proteine die eine Zelle selbst hergestellt hat, präsentiert werden. Dies geschieht in fast jeder Körperzelle auf MHC I Molekülen, und dient dazu, dass durch APZ aktivierte zytotoxische CD8+ T Zellen in Körperzellen hineinsehen können, damit sie diese auf das Vorhandensein eines intrazellulären Virus oder einer maligne Entartung überprüfen können. Wenn eine der CD8+ T Zellen ein fremdes Peptid erkennt, induziert sie den programmierten Zelltod (Apoptose). Auf diese Weise können virenbefallene Zellen oder Tumorzellen von CD8+ T Zellen erkannt und eliminiert werden. Antigenpräsentation an CD4+ T Zellen ist den APZ, zum Beispiel den Dendritischen Zellen (DZ) oder Makrophagen, vorbehalten. Nur sie besitzen die dazu notwendigen MHC II Moleküle, so dass nur sie die Entscheidungsbefugnis zum Einsatz von CD4+ T Helferzellen besitzen. Wir haben uns schwerpunktmäßig mit der Präsentation extrazellulärer Antigene an CD8+ T Zellen befasst, die auch als Kreuzpräsentation (Cross-presentation) bezeichnet wird [1]. Dieser Mechanismus ist zur Aktivierung der CD8+ T Zellen, die einmal virusinfizierte oder maligne entartete Zellen lysieren sollen, notwendig. Denn wenn das Virus die APZ nicht infiziert, müssen sie Antigen von außen aufnehmen, um ihre Aufgabe der Präsentation zu erfüllen. Ähnliches gilt für Tumoren, die nicht aus APZ hervorgehen, also die weitaus größte Zahl der Malignome. Zudem spielt die Kreuzpäsentation eine Rolle in der Aufrechterhaltung der Immuntoleranz gegen unseren eigenen Körper [2]. Wir haben die Frage untersucht, wie eine APZ entscheidet, ob sie ein Antigen mit MHC I oder MHC II präsentiert, also ob sie CD8+ oder CD4+ T Zellen aktiviert. Diese Entscheidung muss gut reguliert werden, denn die Aktivierung der falschen Zellpopulation könnte ineffektiv und im schlimmsten Fall sogar schädlich sein.
Endozytosemechanismen und Antigenpräsentation
In unseren kürzlich publizierten Studien [3, 4] haben wir gezeigt, dass APZ über den sog. Mannose-Rezeptor (MR) ein Modell-Antigen, nämlich OVA (Ovalbumin = Hühnereiweiß) aufnehmen und damit nur CD8+ T Zellen aktivieren können. Mit dem sog. Scavenger Rezeptor (SR) fanden wir einen weiteren Rezeptor zur Aufnahme von OVA. SR-endozytiertes OVA wurde nur an CD4+ T Zellen präsentiert. Schließlich haben wir gezeigt, dass pinozytiertes Antigen ebenfalls nur an CD4+ T Zellen präsentiert wird. Unsere Resultate zeigten, dass der von der APZ gewählte Mechanismus der Antigen-Aufnahme darüber entscheidet, auf welchem Wege das Antigen präsentiert wird. Der Grund dafür ist, dass das Antigen für die Präsentation an CD4+ und CD8+ T Zellen in zwei unterschiedliche Zellorganellen aufgenommen wird. In dem einen wird prozessiertes Antigen nur auf MHC I geladen, in dem anderen nur auf MHC II. Für letzteres war bereits bekannt, dass es sich bei dem betreffenden Organell um späte Endosomes/Lysosomen handelt. Für die Kreuzpräsentation war das zuständige Organell nicht identifiziert. Wir konnten zeigen dass es sich hierbei um ein frühes Endosomales Kompartment handelt, das als„stable early endosomes" bezeichnet wurde. Das bedeutet, dass das Antigen nicht nach der Aufnahme innerhalb der APZ sortiert wird, um zu entscheiden, wie es präsentiert wird, sondern bereits bei der Aufnahme die Art der Präsentation und somit die Art der Immunantwort fest steht.
Eine neue Theorie der Antigenpräsentation
Aus unseren Ergebnissen konnten wir eine neue Theorie der Antigenpräsentation postulieren, in der die Endozytosemechanismen eine zentrale Rolle spielen. Das bisherige Modell sah vor, dass das Antigen von den frühen Endosomen über späte Endosomen in die Lysosomen gelangt, wo es auf MHC II geladen wird, und dass das zu kreuzpräsentierende Antigen vorher aus diesem Pfad abgezweigt werden muss, um auf MHC I geladen zu werden. Dieser intrazelluläre „Kreuzungsmechanismus" konnte jedoch trotz großer Anstrengungen nicht identifiziert werden [5, 6]. Nach unserem Modell (Abb. 1) erfolgt die Sortierung in das Kreuzpräsentationskompartment bereits auf Rezeptor-Ebene [4]. Die Rezeptorabhängigkeit der Kreupräsentation in unserem Modell erlaubt es der APZ, diesen Vorgang durch die Veränderung der Rezeptorexpression zu regulieren. Dies erklärt, warum zur Kreuzpräsentation fähige APZ Populationen durch Expression bestimmter Rezeptoren gekennzeichnet sind.
Neue Möglichkeiten zur Impfstoffentwicklung
Eine wichtige Implikation ergibt sich für die Impfstoffentwicklung gegen Viren oder Tumore, die dem Zweck dienen, spezifische CD8+ T Zellen zu aktivieren [7]. Wenn man einen Impfstoff so modifizierte, dass er über den Mannose-Rezeptor in die Zelle gelangte, würde dadurch die Immunantwort antiviralen bzw. antitumor Charakter besitzen. Hierzu müsste aber sichergestellt werden, dass der Impfstoff auch an CD4+ T Zellen präsentiert wird, damit Helferzellen für die CD8+ T Zellen zur Verfügung stehen. Wenn dies nicht der Fall ist, würde CD8+ T Zell Toleranz resultieren [1, 2]. Dies könnte bei der Therapie von Autoimmunerkrankungen wie z.B. Typ I Diabetes mellitus nützlich sein, da man hier eine Eliminierung autoreaktiver CD8+ T Zellen erzielen muss [8].










